Autores
- ALINE DOS SANTOS ACOSTAUNIVERSIDADE ESTADUAL DE MATO GROSSO DO SULEmail: nyna98@outlook.com
- SIDNEY KUERTENUNIVERSIDADE ESTADUAL DE MATO GROSSO DO SULEmail: sidneykuerten@uems.br
Resumo
O rio Miranda nasce no sudoeste do estado de Mato Grosso do Sul e flui por 756
km, cortando diferentes unidades geológicas até desaguar no rio Paraguai, à
oeste do Estado. Ao longo de todo o seu curso, é possível identificar inúmeras
feições geomorfológicas relictas, dentre as quais os meandros abandonados se
destacam. Neste estudo, foi analisado o conteúdo sedimentar de um desses
meandros abandonados identificados no alto curso fluvial, o qual, a partir de um
testemunho raso de sondagem recuperado (1,75 m). O ponto amostrado em um
paleocanal (21°26’ 38.55” S; 56° 9’ 27.17” O) no município de Jardim (MS). Foram
coletadas amostras do testemunho a cada 5 centímetros, totalizando 35
intervalos, as quais foram processadas para descrição granulométrica e análise
da presença de microfósseis segundo a metodologia proposta por Volkmer-Ribeiro
(1985) para confecção de lâminas microscópicas. Oitenta e sete lâminas foram
produzidas (três lâminas para cada intervalo amostrado).
Palavras chaves
Paleocanal; Espículas de Esponjas; Fitólitos; Pulsos de Inundação; Microfósseis
Introdução
Assim como é importante entendermos o passado da humanidade para explicar o
atual momento da mesma e arriscar palpites sobre o seu futuro, também é de
extrema importância sondarmos a constituição dos aspectos físicos (condições
ambientais) pretéritos para compreender os processos, bem como sua apresentação,
atuais e se possível antever alterações futuras.
Uma das formas de identificar características pretéritas da Terra ocorre através
de estudos paleoambientais. Nesse tipo de investigação são utilizados dados
conhecidos com proxy, ou seja, dados que tem a função de ligar o pesquisador a
seu objeto de estudo tendo como base as características das formas outrora
vivas, agora fossilizadas: eis um elo resistente. Constituem-se então num meio
de se “visualizar” o passado.
Isso ocorre porque cada espécie é adaptada a um tipo de ambiente, logo, podemos
a partir de suas identificações supor a configuração paleoambiental local e
investigar possíveis eventos que ocorreram naquela região (VOLKMER-RIBEIRO E
PAROLIN, 2010). Dentre os dados proxy podemos citar as espículas de espojas, os
fitólitos, as frústulas de diatomáceas, os grãos de pólen e os isótopos de
carbono.
Os proxy mais comumente utilizado em pesquisas paleoambientais são os
polinomorfos (pólens e esporos de plantas vasculares, microalgas, esporos de
fungos, dentre outros), mas em ambientes tropicais úmidos onde o intemperismo
age com mais força sobre as rochas causando oxidação do solo e destruindo a
maior parte do material orgânico é de maior relevância a utilização de dados
proxy formados por opala biogênica, como fitólitos, frústulas de diatomáceas e
espículas de esponjas (STEVAUX, 1994 apud PAROLIN; VOLKMERRIBEIRO; STEVAUX,
2008).
Os primeiros estudos paleoambientais no país foram realizados com sedimentos da
Amazônia mais especificamente na Serra dos Carajás, Pará, iniciado por Martin et
al. (1992) seguindo com Sifeddine et al. (1994), Volkmer-Ribeiro e Turcq (1996)
e outros.
Gradualmente as pesquisas foram executadas nas demais regiões, mas os autores
Kalinovski et al. (2016) ressaltam que esse campo de estudo se concentra na
região Norte e no Rio Grande do Sul, enquanto o Nordeste possui menor número de
relatos espongológicos. O país que presenta maior diversidade espongológica
atualmente é o Brasil, porém vale ressaltar que ainda há muitos lugares
inexplorados e que pesquisas com esse cujo ainda não estão bem difundidas
(KALINOVSKI et al., 2016). Segundo Muricy et al. (2011) e Kalinovski (2015), até
2010 havia 443 espécies de esponjas conhecidas no país, destas 380 pertencem a
Classe Demospongiae sendo 53 espécies de água doce pertencentes a apenas três
famílias: Família Spongillidae Gray, 1867; Família Potamolepidae Brien, 1967;
Família Metaniidae Volkmer-Ribeiro, 1986, incluídas na subordem Spongillina.
De acordo com Volkmer-Ribeiro e Machado (2017), dentre as 53 espécies conhecidas
no Brasil tem ocorrência também no Mato Grosso do Sul. As autoras ainda apontam
que provavelmente existem várias outras espécies no estado visto a grande
riqueza natural trazidas pela existência de duas importantes bacias
hidrográficas que o recortam somada ocorrência de quatro biomas que se misturam
e originam um ambiente rico em biodiversidade: o Pantanal.
Neste trabalho cujo escopo foi utilizar microfósseis, principalmente as
espículas de esponjas, para a reconstituição ambiental de um paleocanal do alto
curso do rio Miranda (afluente do rio Paraguai), localizado no município de
Jardim, MS (Figura 1), outros proxies foram encontrados, e em maior quantidade
os fitólitos. Os resultados obtidos não permitiram uma reconstituição ambiental,
mas auxiliaram sobremaneira a compreensão da dinâmica hidrossedimentar que
ocorreu desde o abandono do meandro até os dias atuais com o processo de
colmatação em andamento.
Material e métodos
O ponto de coleta do material analisado está situado entre as coordenadas 21°
26.582'S e 56°9.462'O (Figura 2), cujo acessa pode ser realizado pela estrada
rural que é um prolongamento da avenida Mato Grosso (área urbana de Jardim,
sentido norte) em direção ao ponto turístico “Cemitério dos Heróis”, distante
4,75 km do acesso à rodovia BR-267 (sentido Porto Murtinho).
O local está adjacente ao canal atual do rio Miranda, distante 190 metros, e em
uma altitude 4 metros acima do nível regular do canal atual (220 m de altitude).
Neste trecho ocorrem rochas do Grupo Itararé, Formação Aquidauana, constituída
localmente por conglomerados e arenitos vermelhos à róseos de granulometria
média a grossa, cujas idades do grupo abrange o Carbonífero Superior ao Permiano
Inferior (CPRM, 2006). De acordo com Ross (2011), a área encontra-se na
depressão do Miranda, limitada à leste pelas escarpas da borda da Bacia do
Paraná e a oeste pelas cristas residuais do planalto da Bodoquena.
No local, foi realizada uma sondagem rasa com auxílio de vibrotestemunhador
motorizado. De posse do testemunho de sondagem com 1,75 m de comprimento,
aberto em laboratório, o conteúdo sedimentar foi inicialmente analisado para
descrição das caraterísticas sedimentares presentes (textura do material,
organização e espessura de fácies, presença e tipo de estruturas sedimentares
primárias). A análise do conteúdo proxy presente nos sedimentos foi realizada a
partir da confecção de lâminas (Quadro 01) em um intervalo de 5 cm,
posteriormente visualizados com auxílio do microscópio para descobrimos o
potencial fossilífero do material recuperado.
Para a produção das lâminas o pesquisador deve utilizar equipamentos de proteção
individual adequados ao manuseio dos materiais listados. Após organizar todos os
itens, seleciona-se 1 grama de sedimento colhido do núcleo do testemunho a cada
5 cm. A escolha por coletar de 5 em 5 cm ocorreu a partir da análise das
características sedimentares, cuja variação no testemunho estudado é pouco
expressiva.
Dada a presença de torrões de areia, o material foi destorroado para otimizar a
ação do ácido de forma que ele não estoure muito (reação ocasional com torrões
ou presença de quartzo). Em seguida, as amostras foram transferidas para tubos
de ensaio devidamente identificados, onde foi acrescentado o ácido Nítrico 65%
até atingir 1/3 do tamanho do tubo ou até cobrir a amostra.
Com o auxílio da capela, da lamparina e do pegador de madeira o material foi
queimado por aproximadamente 3 minutos ou até que a fumaça resultante da queima
ficasse clara, evidenciando que toda a matéria orgânica da amostra foi
eliminada. Foi necessário acrescentar às amostras uma gota de Peróxido de
Hidrogênio para que as lâminas não ficassem escuras, pois elas estavam
estourando muito. Posteriormente, as amostras foram lavadas várias vezes com
água destilada e álcool em centrifugação para neutralizar o ácido.
Para montar as lâminas, agita-se levemente o material de forma que as partículas
muito finas fiquem em suspensão e seja possível pipetar amostra de sua parte
mais superficial. O material também pode ser coletado do fundo e destinado à
outra lâmina. Sempre é necessário utilizar material descartável para que as
amostras não sejam contaminadas e lavar com água corrente e álcool os materiais
não descartáveis.
As lâminas foram identificadas e preparadas com 50 μl de amostra processada,
coletadas com pipeta automática (25 μl). Após a secagem da lâmina, o material
foi coberto com Verniz Vitral com auxílio do bastão de vidro e lamínula. Para
maior conservação da lâmina, é indicado deixar a amostra secar por sete dias
para depois analisá-las.
Esses dados foram utilizados para a construção de uma planilha, que foi
processada no Tilia®, software de bioestatística voltado para estudos
paleontológicos, em especial de microfósseis, para gerar um gráfico com a
distribuição dos próxies na coluna sedimentar.
Resultado e discussão
O conteúdo sedimentar recuperado pelo testemunho de sondagem é caracterizado
pela presença predominante de areias finas a muito finas de cor marrom
claro/ocre, com variações entre 127-118 cm e 88-75 cm para areias fina superior
e pequena fração de areia média inferior (Figura 03). Ocorrem clastos e lâminas
de lama (silte, argila, areia muito fina – detalhes I e II na Figura 03) e
fragmentos de carvão nas profundidades de 146, 140, 135 e 37 centímetros.
Os sedimentos observados permitem reconhecer um ambiente com pouca energia,
caracterizado pela presença de sedimentos muito finos (areia muito fina e lama).
Pulsos de inundações podem ser discretamente observados pela presença de areias
fina superior entre 127-118 cm e 88-75 cm seguidos pela granodecrescência
ascendente marcada por lama. O testemunho apresenta em sua maior parte estrutura
deposicional maciça, com poucas marcas de fluxos/processos de sedimentação, fato
que permite associar a inexistência de fluxos de corrente (ou a não preservação
dessas estruturas) e, por conseguinte a caraterização de um ambiente com pouca
ou nenhuma energia de fluxo, muito semelhante às condições hidrológicas atuais
(lago de meandro abandonado em processo de colmatação).
Da amostragem nos intervalos coletados, resultaram 35 pontos ao longo do
testemunho os quais permitiu produzir 87 lâminas (três lâminas para cada
intervalo amostrado). Um total de 136h foram utilizadas para análise das lâminas
e contabilização do conteúdo proxy.
De modo geral, havia um expectativa de encontrar maior quantidade de espículas
de esponjas ao longo do testemunho, entretanto, poucas foram contabilizadas e a
maior parte dos achados foram fragmentos que não permitiram a identificação em
nível de família. Os fitólitos foram predominantes e estão distribuídos ao longo
de todo o testemunho. Ao todo foram contabilizados 227 fragmentos de espículas e
3 megascleras inteiras e 4.829 fitólitos inteiros e 8.709 fragmentos.
Em 158 cm apareceram as formas globular echinate e globular granulate. Em 173 cm
foram encontradas as formas globular echinate e Cylindric sulcate tracheid. A
maior concentração de fitólitos ocorreu entre 128 e 148 cm. Entre 28 e 43 cm a
concentração de fitólitos diminui e ocorre a forma Cylindric sulcate tracheid.
De 18 a 3 cm a quantidade de fitólitos manteve-se baixa com ocorrência de
Bilobate.
Não foram encontrados dados suficientes para basear uma reconstituição em nível
de vegetação, pois um mesmo morfotipo fitolítico pode ser produzido por várias
espécies, sendo necessária a análise de assembleias fitolíticas que permitiriam
a reconstituição florística local. (PIPERNO, 1988, 1991 apud RASBOLD et al.,
2016; LUZ et al., 2015; KALINOVSKI, 2015). Não obstante, ressalta-se que estudos
sobre as morfologias dos fitólitos apresentados pelas plantas modernas são
escassos no Mato Grosso Sul, dificultando a identificação e classificação dos
mesmos. (LUZ, et al., 2015).
Este nível de análise exige um debruçar maior sobre as relações de tipologias
fitolíticas e espécies de plantas produtoras deles, não havendo tempo hábil para
tanto nesse estudo devido ao curto espaço temporal voltado para seu
desenvolvimento. Entretanto, essa investigação poderá ser alvo de estudos
futuros, configurando um amplo campo de análise, visto a escassez destes estudos
no estado de Mato Grosso do Sul.
Em relação às espículas, as megascleras inteiras foram encontradas nas
profundidades de 18, 143 e 158 cm. Como não foram encontradas gemoscleras,
chega-se à conclusão de não houve condições favoráveis para o desenvolvimento de
esponjas in situ. Cabe ressaltar que é característico do rio Miranda apresentar
uma coloração turva ao longo do ano, indicando que há uma considerável carga de
sedimentos transportados. Em contraponto, sabe-se que o ambiente que abriga as
esponjas não pode ter muita carga em suspensão, pois os sedimentos podem
obstruir seus poros e ou mesmo desfazer as esponjas (VOLKMER-RIBEIRO; PAULS,
2000; VOLKMER-RIBEIRO; PAROLIN, 2010).
Também vale ressaltar que, devido às gêmulas, que carregam células de esponjas
em estágio embrionário, se houver uma de mudança de ambiente as espículas são
capazes de alterar sua função e se adequar às novas características de fluxo
(VOLKMER-RIBEIRO; PAROLIN, 2010). Diante disso e a não ocorrência de gemoscleras
e gêmulas, é sugerida a hipótese de que as espículas inteiras e fragmentos
encontrados no testemunho analisado são alóctones, pois se houvesse esponjas
habitando o canal, elas responderiam a uma alteração no ambiente aquático e
haveria um registro proporcionado pelo aparecimento das gêmulas e gemoscleras.
Por análise do conteúdo de microfósseis e dados estratigráficos, nota-se que ao
ocorrer um pico de fragmentos de espículas e espículas inteiras, ocorre a
drástica redução ou inexistência de fitólitos (inteiros).
Levando em consideração que o registro incompleto dos fósseis nos remete a
processos de remoção/remobilização das camadas deposicionais (SALGADO-LABOREAU,
1998) a ausência de fitólitos foi interpretada como resultado da ocorrência de
fluxo de inundação com energia suficiente para depositar material alóctone ao
lago de meandro abandonado (Figura 04), interrompendo assim a deposição de
fitólitos. Este padrão observado ao longo do testemunho analisado denominamos
neste estudo como pulsos de inundação, destacados na figura 4.
Portanto, a hipótese aventada é a de que esses intervalos representam picos de
inundação com a reativação do fluxo no meandro abandonado, carregando o material
superficial que estava sendo depositado durante a fase de estabilidade (com
maior quantidade de fitólitos) pela vegetação circunvizinha e, por conseguinte,
com a redução do fluxo, a deposição de sedimentos contendo as espículas
fragmentadas e raras megascleras inteiras. Sendo que a própria fragmentação da
maior parte das espículas encontradas sugere essa turbulência ou aumento da
força do fluxo típico de ambiente lótico. Também é possível supor que os
intervalos que apresentam queda na quantidade de fitólitos inteiros representam
inundações de menor porte, cuja força não foi suficiente para que houvesse um
grande carreamento desse material depositado no lago.
Partindo do princípio do atualismo podemos salientar que as fases de
estabilidade hidrológica do lago estudado remetem a períodos pós-inundação,
período em que os sedimentos finos tendem à paulatina decantação concomitante a
entrada de fitólitos da vegetação presente. Fluxos superficiais decorrentes de
precipitações também são aventados para explicar a disposição de areias finas ao
longo do testemunho de sondagem, carreadas para a lagoa. Na figura 05 é possível
observar a condição do lago em 2018 com típica coloração após episódio de
reativação do canal por fluxo de inundação do rio Miranda.
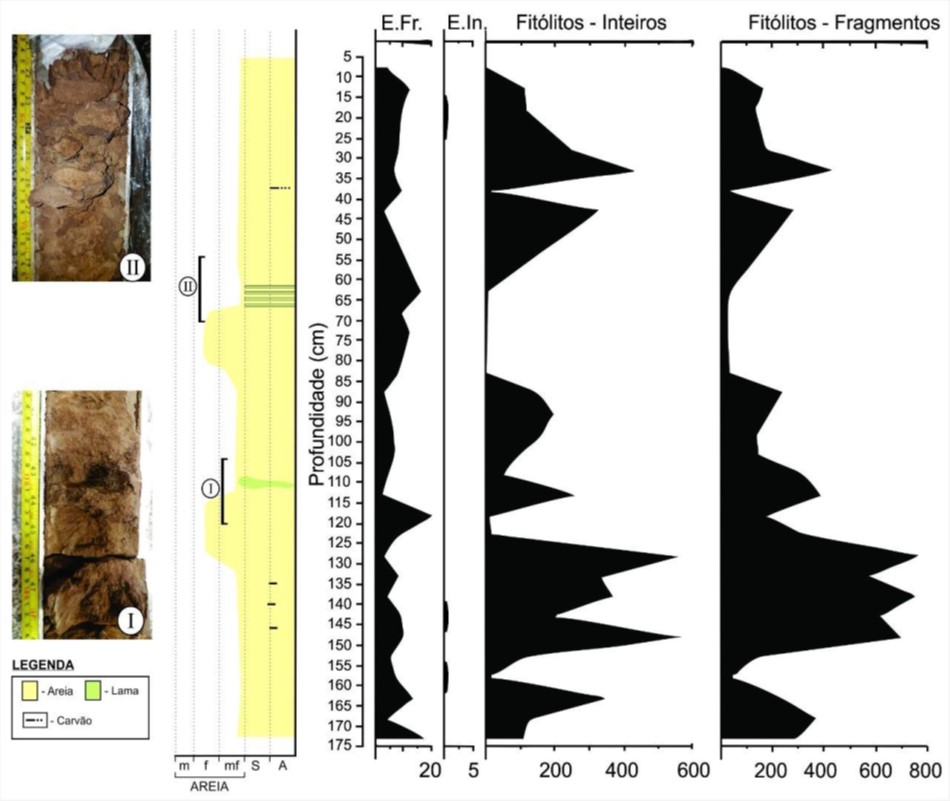
Gráfico de ocorrência de microfósseis nos intervalos amostrados do testemunho de sondagem e perfil estratigráfico.

Imagem aérea do lago estudado. A coloração do lago é resultado da entrada de fluxo de inundação causado pelo rio Miranda no final de outubro de 2018.
Considerações Finais
Através dessa pesquisa foi possível inferir que o registro estratigráfico não
apresentou típicas fácies fluviais. Provavelmente, este registro poderá ser
encontrado em profundidade maior que aquele que foi obtido pelo testemunho. O
registro fóssil observado também revelou que o lago existente não apresentou
condições ambientais favoráveis ao desenvolvimento de uma espongofauna,
provavelmente pela elevada e frequente turbidez da água.
Contudo, os resultados foram satisfatórios e intrigantes; é inevitável
questionar quando ocorreram esses eventos de inundação e quais fitofisionomias
os fitólitos revelariam para o período abrangido pelo testemunho e, afinal,
quanto tempo foi necessário para que ocorresse a deposição de 175 cm de
sedimentos (datações absolutas). Diante disso, salienta-se a necessidade de
continuidade desse estudo para maior detalhamento da análise paleoambiental.
Através da correlação dos resultados da quantificação dos microfósseis e
descrição do perfil sedimentar foi possível realizar uma análise do
comportamento deposicional do canal e a partir deste do seu fluxo, caracterizado
por esse estudo como sendo de baixa energia, visto que a presença de materiais
finos, silte e argila, predominaram em todo testemunho.
Portanto, evidencia-se a validade do método para interpretação paleohidrológicas
local, visto que foi possível realizar esta correlação mesmo sem a identificação
das espécies de esponjas e o grupo de famílias que deram origem aos fitólitos
presentes. A datação de sedimentos deste testemunho e a recuperação de material
de outros meandros abandonados, poderão auxiliar na compreensão da dinâmica das
mudanças fluviais que o canal do Miranda evidencia ao longo de todo o seu curso,
cujos fatores associados podem incluir mudanças ambientais e estruturais ao
longo do sistema fluvial integrado ao rio Paraguai.
Agradecimentos
Aproveitamos para formalizar os agradecimentos à Universidade Estadual de Mato
Grosso do Sul, sem a qual não haveria tantas descobertas e a realização dessa
pesquisa não seria possível.
Referências
CHRISTOPHERSON, R. W. Geossistemas: uma introdução à geografia física. 7ª edição. Bookman: Porto Alegre, 2012.
KALINOVSKI, E. C. Z. PALEOAMBIENTES QUATERNÁRIOS DA PLANÍCIE DO RIO IAPÓ, CASTRO, PARANÁ. (Dissertação de mestrado) Universidade Estadual de Maringá: Paraná, 2015.
KALINOVSKI, E. C. Z; PAROLIN, M.; SOUZA FILHO, E. E. Esponjas de água doce na América do Sul: o estado da arte da produção científica no Brasil: o estado da arte da produção científica no Brasil. In: Terræ Didatica, 2016. Disponível em: <http://www. ige.unicamp.br/terraedidatica/>
LUZ L.D., KALINOVSKI E.C.Z., PAROLIN M., SOUZA FO E.E. Estágio Atual do Conhecimento sobre Fitólitos no Brasil. Terræ Didatica. 2015. Disponível em: <http://www. ige.unicamp.br/terraedidatica/>.
PAROLIN, M.; VOLKMER-RIBEIRO, C.; STEVAUX, J. C. USE OF SPONGOFACIES AS A PROXY FOR RIVER-LAKE PALEOHYDROLOGY IN QUATERNARY DEPOSITS OF CENTRAL-WESTERN BRAZIL. In: REVISTA BRASILEIRA DE PALEONTOLOGIA, 2008.
RASBLD, G. G. PAROLIN, M. CAXAMBU, M. G. RECONSTRUÇÃO PALEOAMBIENTAL DE UM DEPÓSITO SEDIMENTAR POR ANÁLISES MULTIPROXY, TURVO, ESTADO DO PARANÁ, BRASIL. In: REVISTA BRASILEIRA DE PALEONTOLOGIA, 2016.
RAUGUST, T. A evolução sob a perspectiva do registro fóssil. In: A paleontologia em sala de aula. 1° edição. Editora: Imprensa livre, Porto Alegre, 2015. Cap. 6.
SALGADO-LABOREAU, M. L. A HISTÓRIA ECOLÓGICA DA TERRA. 2ª edição. Editor Edgard Blucher ltda: São Paulo, 1998. Cap. 1 e 2.
SIFEDDINE, A; CHIESSI, C.M.; CRUZ JÚNIOR, F. W. INFORMAÇÕES PALEOCLIMÁTICAS BRASILEIRAS. In: BASES CIENTÍFICAS DAS MUDANÇAS CLIMÁTICAS: PRIMEIRO RELATÓRIO DE AVALIAÇÃO NACIONAL. COPPE. Universidade Federal do Rio de Janeiro, Rio de Janeiro, 2014. Cap. 4. Disponível em: http://www.pbmc.coppe.ufrj.br/documentos_publicos/GT1/GT1_volume_completo_cap4.pdf.
VOLKMER-RIBEIRO; MACHADO. Checklist das esponjas do Estado do Mato Grosso do Sul, Brasil. Iheringia: Série Zoologia. Museu de Ciências Naturais: Rio Grande do Sul, 2017.
VOLKMER-RIBEIRO, C.; PAROLIN, M. As Esponjas. In: PAROLIN, M.; VOLKMER-RIBEIRO, C.; LEANDRINI, J. A. (Organizadores) Abordagem ambiental interdisciplinar em bacias hidrográficas no Estado do Paraná. Campo Mourão; Editora da Fecilcam, 2010. Cap. 4, pág. 105-130.
VOLKMER-RIBEIRO, C.; PAULS, S. M. ESPOJAS DE AGUA DULCE (PORIFERA, DEMOSPONGEAE) DE VENEZUELA. In: ACTA BIOLOGICA VENEZUELICA, VOL 20, 2000.
ZERFASS, G. S. A. A PALEOECOLOFIA ATRAVÉS DOS MICROFÓSSEIS. In: A paleontologia em sala de aula. 1° edição. Editora: Imprensa livre, Porto Alegre, 2015. Cap. 15.

















